わたしはタンガニイカ湖シクリッドの多様性創出・維持機構の解明を目的に研究してきました(2〜9)。 また国内では、カワウの食性を研究してきました(1)。
| 9. | オス色彩二型維持機構の解明(進行中) |
| 8. | 見えない「夫婦の絆」の検出 |
| 7. | 体サイズを抑える自然選択 |
| 6. | 産卵の月周期 |
| 5. | 分断自然選択 |
| 4. | 異型交配 |
| 3. | 種同定システム |
| 2. | タンガニイカ湖シクリッドの分類・系統分類 |
| 1. | カワウの食性 |
8.見えない「夫婦の絆」の検出
|
「夫婦の絆」に注目することは、動物の社会構造を解明する上で重要です。
一般に、ペアを作る生物ではメスとオスが一緒に行動するため、どのメスとどのオスがペアを作っているのか、見た目で分かります。
しかし、X. rotundiventralisは水中に高密度の群れを作るため、見た目だけではペアが維持されているのかどうか分かりません。
そこで、分子生物学的手法を用いました。
この魚は、メスとオスの両方が口内で子供を育てます(口内保育)。 そして、メスの口には卵から大きな稚魚まで、さまざまなサイズの子供が入っているのに対し、オスの口には大きな稚魚しか入っていません。 このことから、最初はメスが口内で卵や小さな稚魚を育て、稚魚がある程度大きくなってからオスに渡すことが分かっていました(Yanagisawa et al. 1996)。 しかし、オスが自分の交配相手から子供を受け取っているのか、つまり、オスが自分の本当の子供を保育しているのかは、分かっていませんでした。 そこで本研究では、口内保育をしている大人の個体と、その口の中の子供の親子関係をDNA判定しました。 すると、メスもオスも、口内の子供の本当の親であることが分かりました。 このことから、メスは自分が生んだ子供を口内保育し、オスは、自分が受精させた子供を交配相手のメスから受け取っていることが分かりました。 このことから、少なくとも、産卵から子供の受け渡しまでの期間、夫婦の絆が維持されていることが分かりました。 本研究は、見えない「夫婦の絆」を検出した初めての研究です。 また、「夫婦の絆」を維持する魚は普通、2匹だけの縄張りを作って、他個体の侵入を拒みます(一夫一妻)。 多くの夫婦が集まって群れを作る魚の発見は今回が初めてで、とても珍しい現象と考えられます。 Takahashi T et al. (published online before print) Biology Letters |
 Xenotilapia rotundiventralisの群れです 左上や中央上の口が膨らんだ個体は口内保育中のようです |
7.体サイズを抑える自然選択
|
動物の体サイズは、大きい方が有利となる選択と小さい方が有利となる選択のバランスで決まると考えられます。
オスに対する性選択(sexual selection)とメスに対する繁殖力選択(fecundity selection)は、それぞれ大きな体サイズを進化させる選択圧として一般に認識されています。
しかし、小さな体サイズを進化させる選択圧については、あまりよく知られていません。
体サイズを制限する選択圧には、一般に内的な要因(生理的制約など)が関係していると考えられます。
しかし、外的な環境要因によって体サイズが制限される例は、あまり知られていません。
そこで本研究では、貝を隠れ場所や産卵場所として利用するTelmatochromis temporalisの矮小型について、体サイズを決定する要因を調べました。
この魚は、オスの方がメスより大きい性的サイズ二型を示します。 フィールド観察から、オスは縄張りを形成し、体サイズが大きいほど、縄張り内のメスの数が多い傾向にあることがわかりました。 このことは、大きなオスが有利となる性選択が働いていることを示唆します。 しかしフィールドでの移植実験から、大きくなりすぎると(体長40mm以上)、貝に隠れられなくなり、生存に不利となることが示唆されました。 一方メスでは、大きな個体ほど卵巣内に多くの卵を持っていたので、大きなメスほど有利となる繁殖力選択が働いているものと考えられます。 しかし、メスは貝の中のとても深く、とても奥の方に産卵します。 もしかしたら、貝の奥の狭い空間に産卵することにより、卵に対する補食圧を減らしているのかもしれません。 メスの体サイズは、大きいと多くの卵を産めるが狭い安全なところには産めない、逆に小さいと狭い安全なところに産卵できるが産卵数が少なくなる、というトレードオフ関係にあるのかもしれません。 本研究では、オスでは成魚への補食圧が体サイズを抑える自然選択の要因となり、メスでは卵への補食圧が体サイズを抑える自然選択の主な要因となっていることが示唆されました。 このように、T. temporalisの矮小型では、雌雄ともに自然選択で体サイズが制限され、しかもその選択圧が雌雄で異なることが示唆されました。 これまで動物においてこのような例は知られておらず、初めての報告となります。 Takahashi T et al. (2012) Hydrobiologia 684:35-44 |
 左の大きい魚がオス、右の小さな魚がメスです それぞれ別の貝を利用します 同じ貝に一緒に入ることは、とても稀です 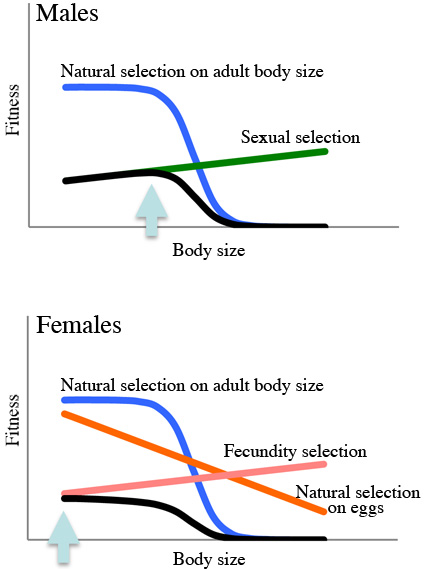 フィットネスのイメージ図 オスの方が、フィットネスが最大になる体サイズが大きいです |
6.産卵の月周期
|
多くの淡水魚は産卵が月周期に同調しませんが、タンガニイカ湖シクリッドでは同調します。
その説明のひとつとして、肉食性のナマズ類が新月の夜に活発になるため、この時期に繁殖するのを避けていると考えられます。
本研究では、分断自然選択で紹介したテンポラリスの普通型と矮小型の産卵周期を調べました。 すると、普通型では月周期産卵が検出されましたが、矮小型では検出されませんでした。 普通型は岩の下に産卵しますが、矮小型は貝の奥で産卵します。 貝の奥は狭く、卵や稚魚への捕食を避けるのに効果的であると考えられます。 矮小型は、産卵を月周期に同調させなくても、安全に子供を育てられるのかも知れません。 Takahashi T (2010) Hydrobiologia 644:139-143 |
 矮小型は貝のとても奥に産卵します(貝を割って中を見易くしています 割る前は殻高4 cmくらい) |
5.分断自然選択
|
分断自然選択は、異なる環境への適応によって、集団間に異なる形態や生態が進化する機構です。
この機構は多くの種分化に関与してきたと考えられますが、意外にも明確に示した例は限られています。
この解明を難しくしている理由として、多くの種では進化が大昔に完了してしまったこと、そして一般にひとつの進化現象に複数の要因が複雑に関係してきたことが挙げられます。
そこで私は、最近起きた、単純な進化を調べるため、タンガニイカ湖に生息するテンポラリス(Telmatochromis temporalis)という魚を調べました。
この魚には岩の隙間を隠れ場所とする体長8センチほどの普通型と、巻貝の空殻を隠れ場所とする4センチほどの矮小型がいます。 これらを分子生物学的に調べたところ、湖南部の二カ所で、矮小型が極めて最近、普通型から繰り返し進化したことが分かりました。 普通型と矮小型には隠れ場所以外に目立った違いがないため、矮小型の小さな体は、貝に隠れることに適応した結果であると考えられます。 これは、分断自然選択の明確な例であるといえます。 Takahashi T et al. (2009) Mol Ecol 18:3110-3119 Takahashi T (2004) J Fish Biol 65:419-435 |
 左が矮小型で右が普通型 どちらも成熟オスです  矮小型が貝に入っているところ |
4.異型交配
|
ある研究によると、女性は自分と異なるHLAの遺伝子型を持つ男性の匂いを好む傾向があるそうです(例えばSantos et al. 2004)。
これは、異なるHLA型の個体同士がペアになり易いことを示しているのかも知れません。
異なる表現型の個体同士がペアになる傾向を「異型交配(異類交配)」といいます。
先ほどのヒトの例では、HLAにおいてヘテロの子供の割合を多くしたり、近親交配を避ける働きがあると考えられています。
本研究では、タンガニイカ湖に生息するペリソダス(Perissodus microlepis)という魚を採集し、口の曲がる向きを調べました。 すると、口の向きが異なる個体同士が高い割合でペアになることを発見しました。 これは、野生集団で異型交配を明確に示した初めての例です。 右向き個体と左向き個体のペアからは、右向きの子供と左向きの子供が1:1に近い割合で生まれることが知られています。 このため異型交配は、集団内での右向き個体と左向き個体を1:1に維持する機構であると考えられます。 しかしこの例では、ヒトの匂いの例と違い、異型交配が近親交配を避ける働きはないようです。 つまり、異型交配ペアの子供が右向き:左向き=1:1で集団中の割合とほぼ同じなので、子供は自分の兄弟と他人を見分けることができないからです。 では、どうして異型交配を行うのでしょうか? それを解明することは今後の課題です。 Takahashi T and Hori M (2008) Biol Lett 4:497-499 |
 子育て中のペリソダスのペア 下がオスで上がメス |
3.種同定システム
|
タンガニイカ湖シクリッドの種同定には経験が必要で、初学者には難しい場合もあります。
そこで、種同定を補助するソフトISTC(科博・MRAC)を作りました。
ISTCでは、文献情報もデータベースとして取り入れられるように工夫しました。 一般に文献に記載された形態情報には形質間の相関が示されていません。 また多くの文献では、形質のレンジ(最小値と最大値)だけが示されています。 このようなデータは、そのままでは統計的に扱うことができません。 ISTCではこのようなデータから各形質の平均と分散を推定し、形質間の相関を無視した判別分析を行いました。 このように簡略化した方法は、解析の精度を下げると考えられます。 しかし実際にテストしてみると、一般の判別分析と比べ、同定精度の低下はほとんど認められませんでした。 このことは、少なくともシクリッドでは、この方法が種同定に有効であることを示しています。 もしこの方法が他の魚種グループでも有効であれば、文献情報を使えるので、少ない労力で判別分析ができるメリットがあります。 Takahashi T et al. (2007) Ichthyol Res 54:399-403 |
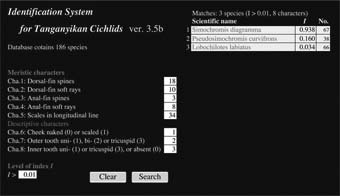 ISTCの画面 |
2.タンガニイカ湖シクリッドの分類・系統分類
|
アフリカ大地溝帯の古代湖であるタンガニイカ湖には約200種類ものシクリッド魚類(カワスズメ科 Cichlidae)が生息し、そのほとんどが湖に固有です。
このためこの魚は、湖内で爆発的に種分化したと考えられ、適応放散のモデルとして世界中の注目を集めています。
しかし、基本となる分類学的研究は十分ではありません。
このことから、新種の発表や族(亜科と属の間の分類階級)の整理などを行いました。
Takahashi T (2008) J Fish Biol 72:603-613 Takahashi T and Hori M (2006) J Fish Biol 68B:174-192 Takahashi T et al. (2006) J Fish Biol 68A:56-67 Takahashi T (2004) Ichthyol Res 51:1-4 Takahashi T and Nakaya K (2003) Copeia 2003:824-832 Takahashi T (2003) Ichthyol Res 50:367-382 Takahashi T (2003) Species Divers 8:1-26 Takahashi T (2003) Ichthyol Res 50:36-47 Takahashi T et al. (2002) Copeia 2002:1029-1036 Takahashi T and Nakaya K (1999) Copeia 1999:101-106 Takahashi T and Nakaya K (1998) Ichthyol Res 45:413-418 Takahashi T and Nakaya K (1997) Ichthyol Res 44:335-346 Takahashi T et al. (1997) Ichthyol Res 44:109-117 |
 タンガニイカ湖 右下の琵琶湖よりずっと大きい 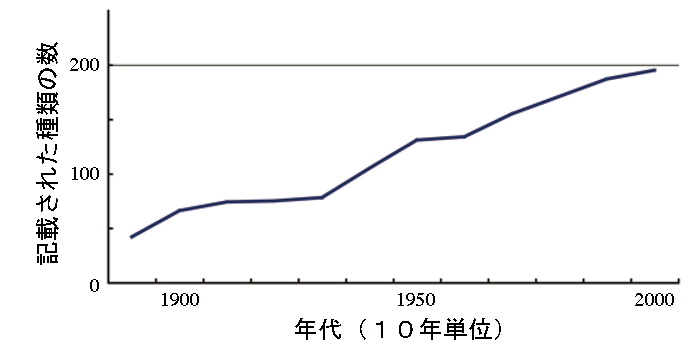 まだまだ新種が見つかりそう・・・ |
1.カワウの食性
|
カワウは近年、琵琶湖周辺で急激に増えた大型の魚食性水鳥で、在来魚への捕食の影響が懸念されています。
そこで、カワウの食性を調べました。
まず、カワウの胃の中で消化された餌魚の骨の一部から、種類と重さを推定する方法を確立しました。 そして、この方法を用いてカワウの胃内容物を魚種ごとに分け、それぞれの総重量を推定しました。 その結果、春夏には在来種で琵琶湖の産業重要種であるアユを、そして秋冬には外来種のオオクチバスを多く食べていることが明らかになりました。 このようなカワウ食性の季節変化は、琵琶湖産アユの生態と関係していると考えられます。 アユは年魚(1年1世代で、産卵してすぐに死ぬ)で、琵琶湖の場合、流入河川で9〜10月くらいに産卵します。 孵化した稚魚は流下し、日中は琵琶湖の底でじっとしているようです。 稚魚の中でも大型の個体は、3月くらいから流入河川に遡上し始めますが、小さな個体は琵琶湖に留まります。 一般に、この早い時期に遡上する大きな個体を「オオアユ」、湖に残る小さな個体を「コアユ」と呼びまが、厳密な区別ではありません。 コアユは6〜7月に、湖の表層近くで「マキ」と呼ばれる高密度の群れを作ります。 今回の調査では、この「マキ」が出現する時期と、カワウの胃からアユが大量に出現する時期がだいたい一致しました。 また、カワウの胃から見つかったアユの体サイズは、(オオアユではなく)コアユと同じくらいでした。 ウの仲間は、群れを作る比較的小さな魚を好んで捕食することが報告されています。 琵琶湖のカワウも、コアユが「マキ」を作る時期には、コアユを優先して食べていたのでしょう。 この食性調査は、1998年の3〜7月および2001年4月〜翌年2月の間に捕獲されたカワウについて行いました。 これらのサンプルから推定されたアユの捕食量は4〜6月に46トン程度、7〜8月には310トン程度でした。 アユの漁期はおおよそ11月から翌6月。 この間の漁獲量は年によって違いますが、千トン前後で推移しています(1998〜2002年)。 これらのことから、カワウによる捕食量は漁獲量より少ないことが示唆されました。 一方、今回の胃内容物調査では、アユ産卵期の9〜10月のデータがほとんどありません。 もしカワウが産卵直前のアユを食べていれば、アユ資源量に対する影響は予想以上かも知れません。 またこの研究では、カワウが冬期間の貴重な餌として、外来魚のオオクチバスを利用していることが示唆されました。 もしかしたら、一時は絶滅の危機にまで瀕した日本のカワウ(在来種)が、外来魚の移入によって個体数を回復し、今度は多くなり過ぎたのかもしれません。 Takahashi T et al. (2006) Fish Sci 72:477-484 高橋ほか (2003) 日本水産学会誌 69:643-645 高橋ほか (2002) 日本水産学会誌 68:576-578 |
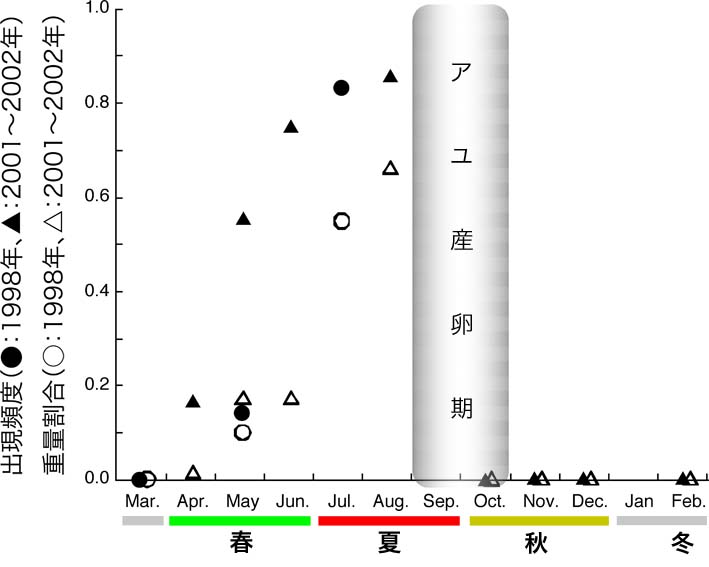 琵琶湖のカワウの胃内容物におけるアユの出現頻度と重量割合 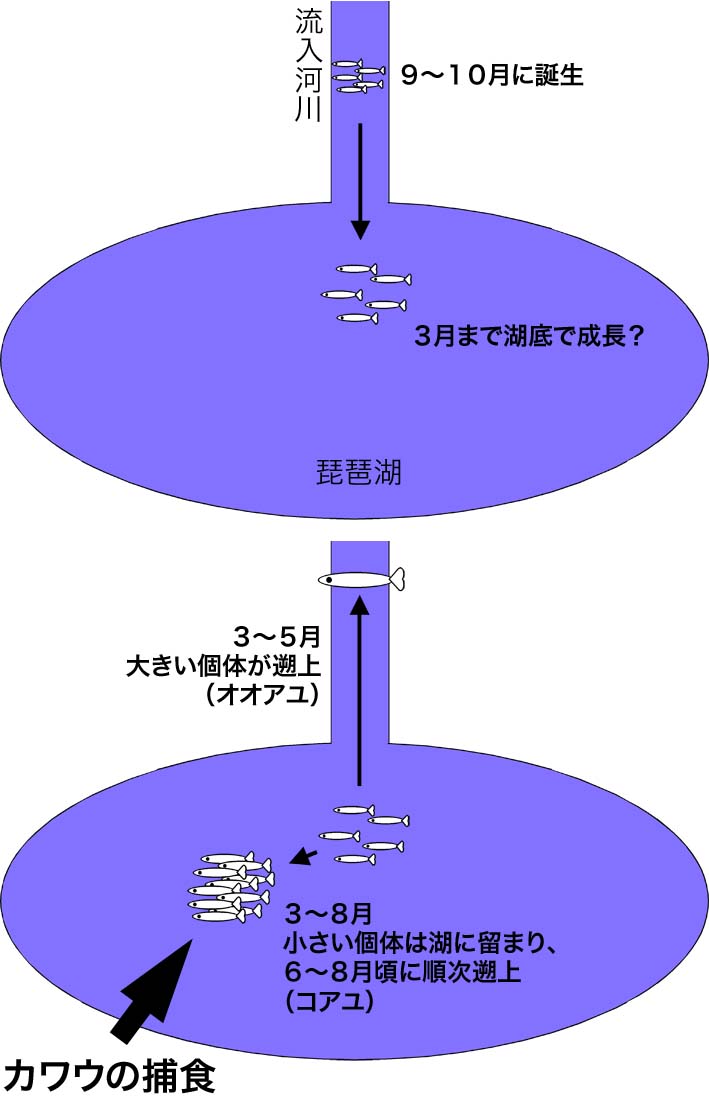 琵琶湖産アユの一生(概念図) |
ホームに戻る